The Team is a combined unit of the Department of
Biotechnology, University of Szeged, and the Laboratory of
Microbial Gas Metabolism at the Institute of Biophysics,
Biological Research Center. In the focus of our investigations
is the molecular understanding of microbial utilization and
generation of hydrogen and methane.
| HYDROGENASE | ||||
| The understanding of the molecular
fundamentals of hydrogen production and utilization among
microbes is a goal of supreme importance both for basic
and applied research applications. The key enzyme in
biological hydrogen metabolism is hydrogenase, which
catalyses the formation or decomposition of the simplest
molecule occurring in biology: hydrogen.
|
||||
| H2
<=> 2H+ + 2 e-
|
||||
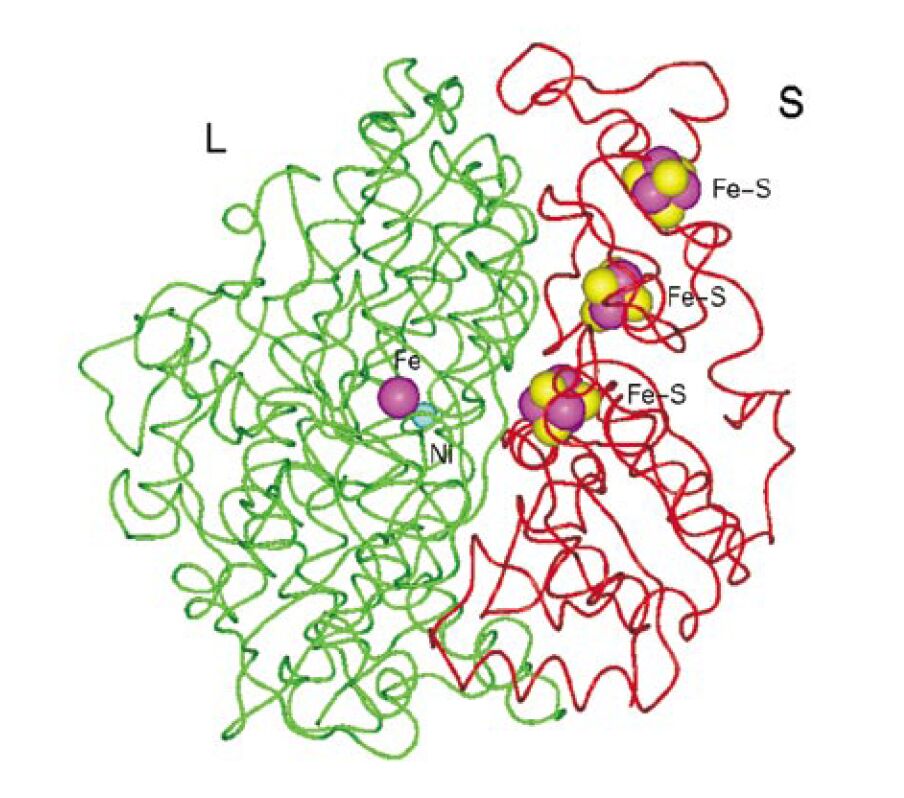
| The simple-looking task is solved by a
sophisticated molecular mechanism. Hydrogenase is a
metalloenzyme, harbouring Ni and Fe atoms. Like most
metalloenzymes, hydrogenases are extremely sensitive to
inactivation by oxygen, high temperature, CO, CN, and
other environmental factors. These properties are not
favourable for the potential biotechnological
applications. In metal containing biological catalysts, it is the protein matrix surrounding the metal centres which provides the unique environment for the Fe and Ni atoms, allowing hydrogenases to function properly, selectively, and effectively. Therefore, the main goal of our basic research is to understand the protein-metal interaction. The problem is not simple to address, as some of the methods for scientific investigation provide information on the metal atoms themselves without directly observing the protein matrix around them. Other modern techniques at our disposal reveal details of the protein core, but do not expose the metal centres within. A combination of these two approaches, i.e., molecular biology and biophysics, is expected to uncover the fine molecular details of the catalytic action of metalloenzymes.
HYPERTHERMOPHILES One of the purple photosynthetic bacteria (Thiocapsa roseopersicina) contains a hydrogenase, which displays outstanding stability among enzymes possessing the same catalytic function. Heat stability does not provide an advantage for the bacterium in its natural, cold-water marine environment. Understanding, through molecular biological studies, the physiological properties and the molecular factors that stabilize this hydrogenase is expected to help design various enzymes equipped with resistance to the inactivating effect of high temperature, oxygen, and/or proteolytic attack for future biotechnological use. Random and site directed deletion mutagenesis studies revealed the presence of three other hydrogenases in this organism, thus, T. roseopersicina contains representatives of each NiFe hydrogenase classes known today. Thus, this bacterium offers unique possibilities to study hydrogenase organisation, assembly and regulation of biosynthesis in a single species. The genome of T. roseopersicina is being sequenced in a collaborative effort and a proteomic approach has been launched in order to understand the complex problem. Hyperthermophilic microorganisms grow above 80oC and cannot multiply below 70oC. By definition, enzymes operating in these unusual creatures are heat stable, therefore, they offer an obvious advantage for biotechnological applications. In their chemolithotroph mode of growth, hydrogen metabolism plays a crucial role. We have developed a novel technique to study their microbiology, molecular biology and genetics. |
||||
| METHANOTROPHS | ||||
| Methanotrophic bacteria typically
contain another metalloenzyme, methane monooxygenase
(MMO). Methanotrophs utilize methane as their sole source
of carbon and energy. Methane is oxidized to CO2
through methanol, formaldehyde and formate
intermediaries. The first and most important step in
this sequence of reactions is the methane to methanol
conversion, catalysed by MMO. MMO can also attack
several compounds representing serious environmental and
public health hazard, such as chlorinated hydrocarbons.
A clear view of the methane and hydrogen metabolism in methanotrophs will be significant for advancing molecular enzymology as well as for practical utilization of methanotrophs to produce alternative energy sources and environmental protection and remediation, improving the general quality of life. |
||||
| BIOHYDROGEN | ||||
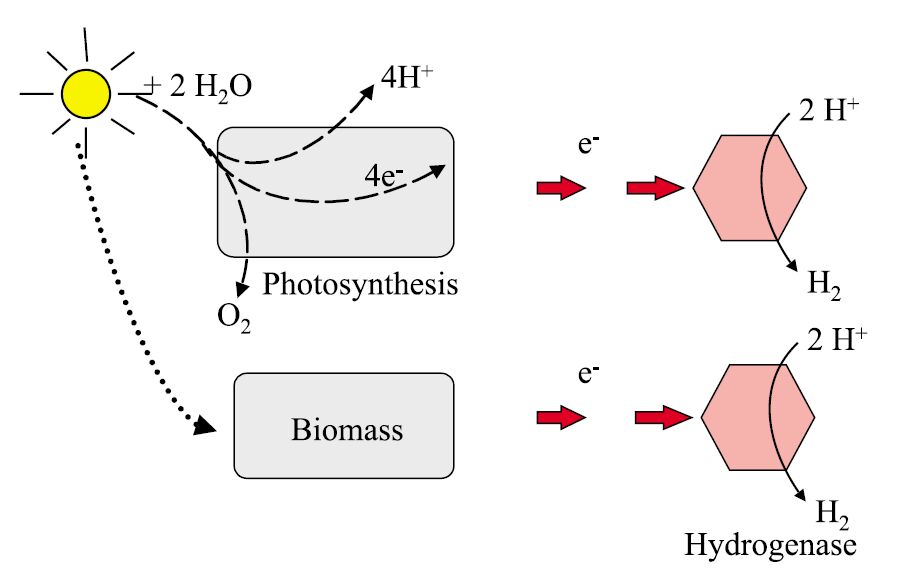
Among the alternative energy carriers, hydrogen is preferred. Hydrogen can be produced in biological processes: solar energy captured by the photosynthetic apparatus is converted into chemical energy through water splitting; the reaction forms oxygen and can also produce hydrogen. Upon utilization, these components are combined to form water, and energy is released in a cycle driven by the practically unlimited and safe energy source of the Sun. In addition to offering an alternative for the global energy crisis, biologically produced hydrogen may also serve as reductant for numerous microbiological activities of environmental significance. A combination of hazardous waste disposal with biohydrogen production has been demonstrated using keratin rich waste. In a two-stage fermentation system keratin has been efficiently converted to hydrogen and microbial biomass. Biogas.This principle has been employed in the complex microbiological series of events that leads to biogas formation. Biogas formation is intensified through the microbiological manipulation of the intermediate hydrogen production steps according to our patented procedure. The method has been tested in field experiments at a landfill site and in a waste water sludge treatment plant. |
||||
Rákhely, G., Zhou, Z. H., Adams, M.W.W. and Kovács, K.L. (1999). Biochemical and molecular characterization of the [NiFe]hydrogenase from the hyperthermophilic Archaeon Thermococcus litoralis. Eur. J. Biochem. 266: 1158-1165.
Fodor, B., Rákhely, G., Kovács, Á.T. and Kovács, K.L. (2001). Transposon mutagenesis in purple sulfur photosynthetic bacteria: Identification of hypF, encoding a protein capable to process [NiFe] hydrogenases in ?, ß and ? subdivision of proteobacteria. Appl. Environ. Microbiol. 67: 2476-2483.
Takács, M., Rákhely, G. and Kovács, K.L. (2001). Molecular characterization and heterologous expression of hypCD, the first two [NiFe] hydrogenase accessory genes of Thermococcus litoralis. Arch. Microbiol. 176: 231-235.
Csáki, R.,Bodrossy, L., Hanczár, T., Murrell, J.C. and Kovács, K.L. (2001). Molecular characterization of a membrane bound hydrogenase in the methanotroph Methylococcus capsulatus (Bath). FEMS Microbiol. Lett. 205: 203-207.
Hanczár, T., Bodrossy, L., Csáki, R., Murrell, J.C. and Kovács, K.L. (2002). Hydrogen driven methane oxidation in Methylococcus capsulatus (Bath). Arch. Microbiol. 177: 167-172.
Kovács, K.L., Fodor, B., Kovács, Á.T., Csanádi, Gy., Maróti, G., Balogh, J., Arvani, S. and Rákhely, G. (2002). Hydrogenases, accessory genes and the regulation of [NiFe] hydrogenase biosynthesis in Thiocapsa roseopersicina. Int. J. of Hydrogen Energy 27: 1463-1469.
Csáki, R., Bodrossy, L., Klem, J., Murrell, J.C. and Kovács, K.L. (2003). Genes involved in the copper-dependent regulation of soluble methane monooxygenase of Methylococcus capsulatus (Bath): cloning, sequencing and mutational analysis. Microbiology 149: 1785-1795.
Kovács, Á.T., Rákhely, G. and Kovács, K.L. (2003). Genes involved in the biosynthesis of photosynthetic pigments in the purple sulfur photosynthetic bacterium Thiocapsa roseopersicina. Appl. Environ. Microbiol. 69: 3093-3102.
Bagyinka, Cs., Ősz, J. and Száraz, S. (2003). Autocatalytic oscillations in the early phase of the photoreduced methyl viologen-initiated fast kinetic reaction of hydrogenase. J. Biol. Chem. 278: 20624-20627.
Maróti, G., Fodor, B.D., Rákhely, G., Kovács, Á.T.,
Arvani, S. and Kovács, K.L. (2003). Accessory proteins
functioning selectively and pleiotropically in the biosynthesis
of [NiFe] hydrogenases in Thiocapsa roseopersicina. Eur.
J. Biochem. 270(10): 2218-27.